关键信息
-
基因名
CD46
- 应用
-
别名
CD46; MIC10; TLX; TRA2; Trophoblast leukocyte common antigen; CD46 Molecule,Complement Regulatory Protein; CD46 Molecule,Complement Regulatory protein
-
种属
Human
-
表达系统
E. coli
-
标签
N-His
-
纯度
Greater than 90% as determined by SDS-PAGE.
-
蛋白编号
P15529
-
表达区间
Val147~Lys285
-
分子量
19kDa
-
内毒素
< 1.0 EU per μg protein as determined by the LAL method.
-
性状
Freeze-dried powder
-
缓冲液
PBS, pH7.4, containing 0.01% SKL, 1mM DTT, 5% Trehalose and Proclin300.
-
复溶方法
Reconstitute in ddH2O to a concentration of 0.1-0.5 mg/mL. Do not vortex.
- 个性化定制
-
稳定性测试
The thermal stability is described by the loss rate. The loss rate was determined by accelerated thermal degradation test, that is, incubate the protein at 37℃ for 48h, and no obvious degradation and precipitation were observed. The loss rate isless than 8% within the expiration date under appropriate storage condition.
-
保存条件 & 期限
Samples are stable for up to twelve months from date of receipt at -20℃ to -80℃. Store it under sterile conditions at -20℃ to -80℃. It is recommended that the protein be aliquoted for optimal storage. Avoid repeated freeze-thaw cycles.
-
运输条件
In general, recombinant proteins are supplied as lyophilized powder and shipped at ambient temperature. For bulk packages, the proteins are provided as frozen liquid and shipped with blue ice, unless otherwise requested by the customer.
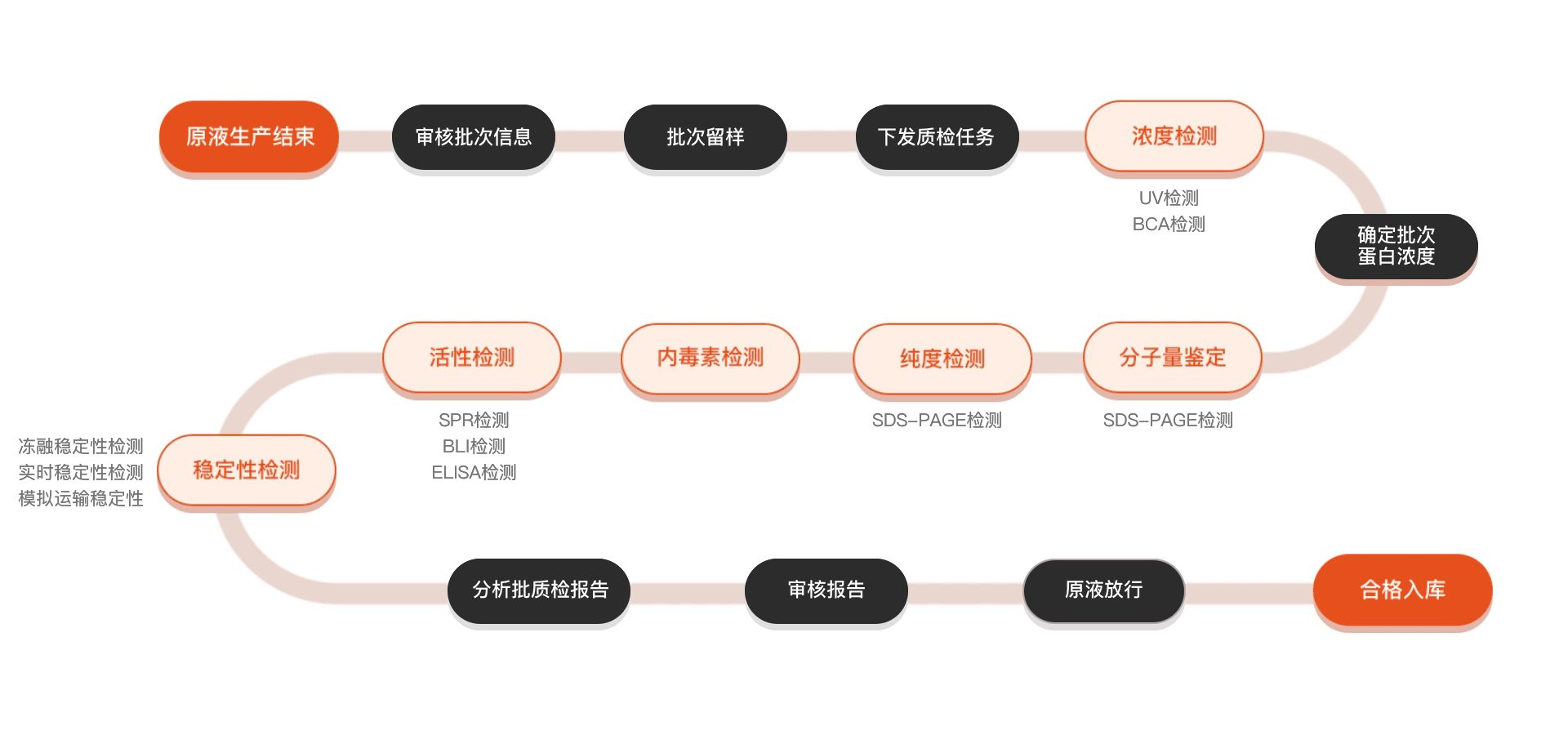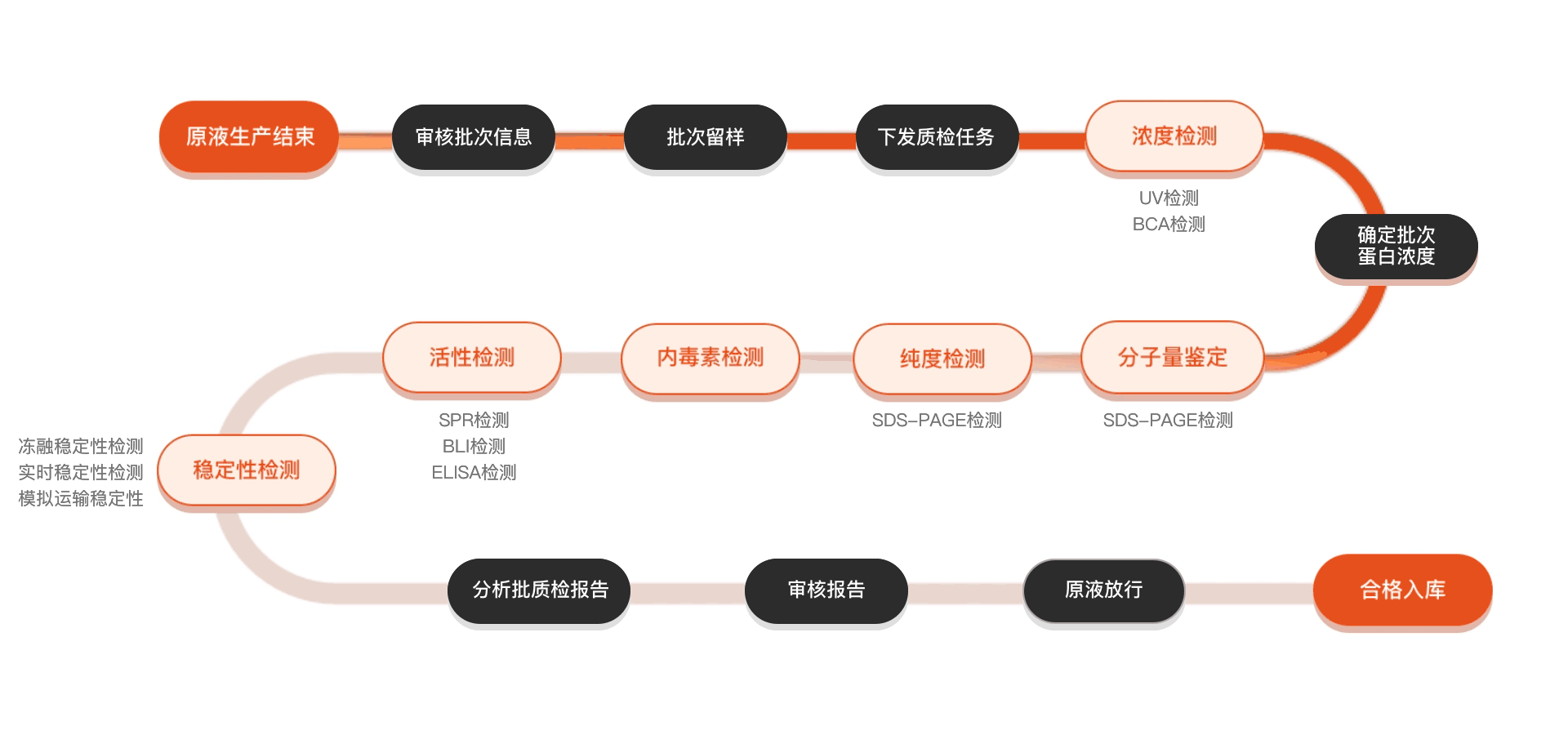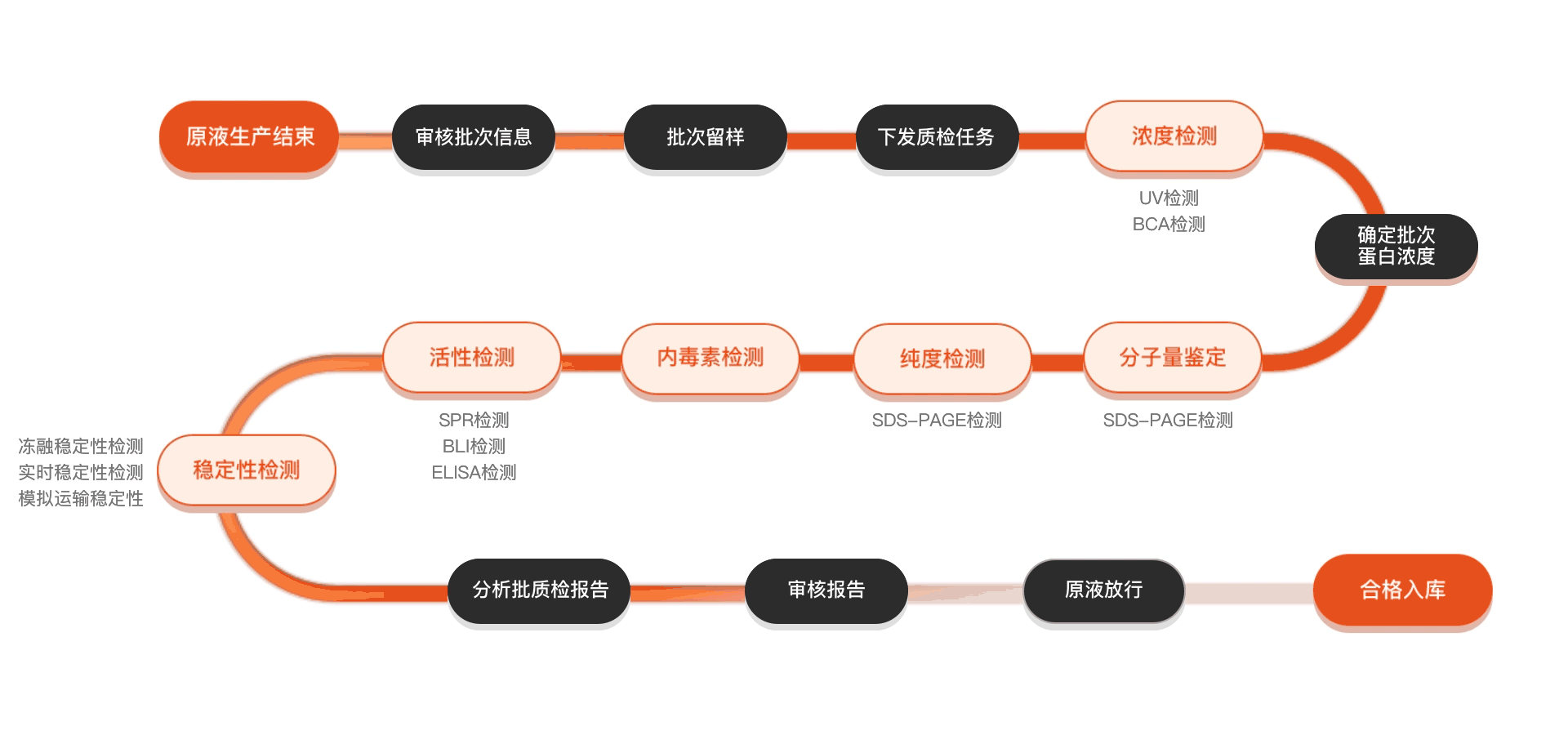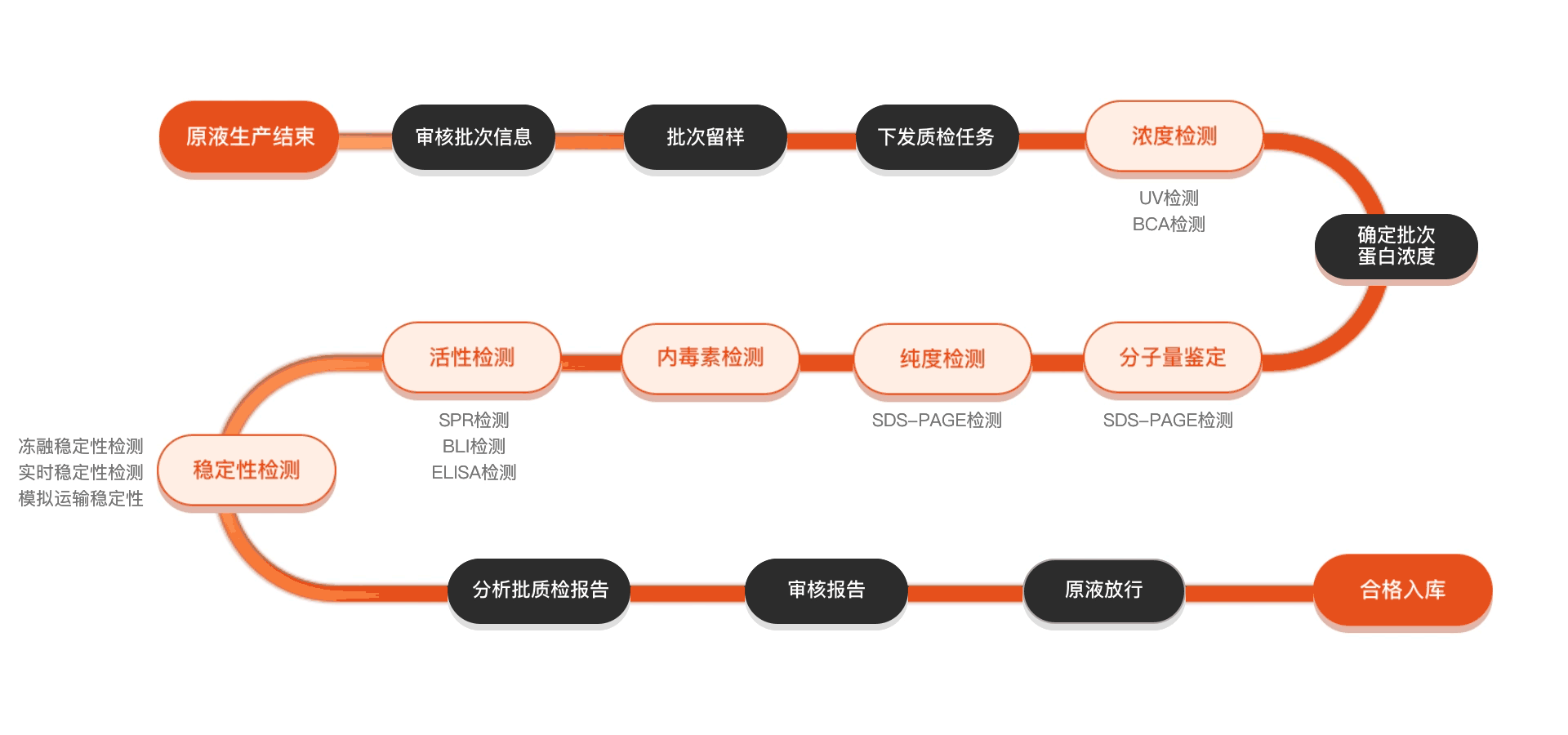
质检流程
相关产品






背景信息
CD46, also known as membrane cofactor protein (MCP), is a crucial regulator of the immune system, primarily recognized for its role in protecting host cells from complement-mediated lysis. It serves as a cofactor for Factor I in cleaving C3b and C4b, thereby modulating the complement pathway and preventing excessive inflammation. Research into CD46 has gained momentum due to its involvement in various pathological conditions, including autoimmunity, infectious diseases, and cancer. The role of CD46 in viral entry, particularly for viruses like the measles and adenoviruses, highlights its significance beyond complement regulation. Recombinant CD46 proteins have emerged as valuable tools for scientific investigation, enabling researchers to study its molecular interactions, expression patterns, and functional implications in health and disease. By generating and characterizing CD46 recombinant proteins, scientists aim to elucidate its mechanisms and explore potential therapeutic interventions, such as enhancing immune responses or developing vaccines. The continued exploration of CD46 and its recombinant forms holds promise for advancing our understanding of immune regulation and addressing various diseases linked to its dysregulation.